Abstract
Are animals’ preferences determined by absolute memories for options (e.g. reward sizes) or by their remembered ranking (better/worse)? The only studies examining this question suggest humans and starlings utilise memories for both absolute and relative information. We show that bumblebees’ learned preferences are based only on memories of ordinal comparisons. A series of experiments showed that after learning to discriminate pairs of different flowers by sucrose concentration, bumblebees preferred flowers (in novel pairings) with (1) higher ranking over equal absolute reward, (2) higher ranking over higher absolute reward, and (3) identical qualitative ranking but different quantitative ranking equally. Bumblebees used absolute information in order to rank different flowers. However, additional experiments revealed that, even when ranking information was absent (i.e. bees learned one flower at a time), memories for absolute information were lost or could no longer be retrieved after at most 1 hr. Our results illuminate a divergent mechanism for bees (compared to starlings and humans) of learned preferences that may have arisen from different adaptations to their natural environment.
Editor's evaluation
This is a very informative and nicely controlled study showing that, when retrieving the value of a food type, bumblebees guide their choices by remembered ranking of feeders instead of focusing on their absolute rewards.
https://doi.org/10.7554/eLife.78525.sa0Introduction
What do animals remember about items out of context? For example, suppose we learn that different options (e.g. coffee shops) result in different reward outcomes (e.g. waiting time and quality), and later we are presented with a choice between two previously encountered options which we have never experienced side-by-side. What types of values do we remember for those options now presented in a novel context? Do our memories of the subjective values for each option contain absolute information (e.g. delay to reward), remembered ranking (how they compared to previous alternatives), or a weighted combination of both?
In typical studies exploring the economic choices of animals including humans, subjects do not have to use distant memories of the options; they are presented with choices where the objective values (e.g. amount, cost, and status) are concurrently visible and can be directly compared. Under such conditions, a wealth of research shows that animals’ choices can be influenced by the presence of additional options (Hunter and Daw, 2021; Spektor et al., 2021). An example of this phenomenon is frequently used in marketing: when given a choice between popcorn options in different sizes, e.g. $3 for small and $7 for large, most people choose the smaller cheaper option, but when a $6 medium option is added, more people choose the large because it now seems like a good deal. Evidence of contextual effects like this on direct assessments has been found across the animal kingdom, e.g. humans and other primates (Berkowitsch et al., 2014; Parrish et al., 2015; Trueblood et al., 2013), bats (Hemingway et al., 2021), birds (Bateson, 2002; Morgan et al., 2012), frogs (Lea and Ryan, 2015), fish (Reding and Cummings, 2017), bees (Shafir et al., 2002), and worms (Iwanir et al., 2019). However, little is known about the type and degree of information (absolute and/or relative) that is encoded in the remembered subjective values of options.
Only more recently have investigations of absolute and relative information traversed into the realm of reinforcement learning, where value must be inferred from memories. Studies on starlings (Pompilio and Kacelnik, 2010) and humans (Bavard et al., 2018; Bavard et al., 2021; Klein et al., 2017) demonstrated that both absolute memories and remembered ranking are combined in particular ways to give rise to these animals’ preferences. So far, however, no other species have been investigated for the roles played by absolute memories and remembered ranking in learned preferences. Here, we examine this in bumblebees (Bombus terrestris), an invertebrate and a key model for examining the economy of decision-making outside of humans (Real, 1996).
Specifically, we adopt an instrumental learning paradigm that combines a contextual training phase and a transfer-test phase (Palminteri and Lebreton, 2021). This paradigm essentially involves two distinct learning contexts (e.g. AB context and CD context; Figure 1), with each context offering two options of contrasting properties (e.g. A>B and C>D in reward sizes). After training, animals’ learned preferences are tested with a novel combination of options (e.g. B vs C). Note that behavioural tests for transitive inference (Guez et al., 2013; Vasconcelos, 2008) involve a similar task design, which includes several training phases and a test phase of new combinations. However, this method provides overlapped relational premises during training (e.g. A>B, B>C, C>D, and D>E) in order to examine if animals can infer the relationship between a pair of options within the chained sequence which had previously not been experienced together (e.g. B vs D). In contrast, our paradigm provides no direct overlapped training between contexts (e.g. A>B and C>D), and therefore, animals cannot infer the relationship between unchained options (e.g. B vs C). Rather, by providing both absolute and relative information during training, our paradigm is used to assess whether, and in what combination, bumblebees retain and utilise absolute and ranking memories.
Figure 1 with 4 supplements

Bumblebees make decisions based on ordinal comparisons.
(A, C, and E) The corresponding sucrose concentration of each stimulus is displayed on a log scale to visually represent their relative differences according to Weber’s Law (Akre and Johnsen, 2014). …
Figure 1—figure supplement 4
Bees can discriminate flowers of different colours and sucrose concentrations.
To ensure that bees were able to learn to discriminate between two differently coloured flowers in our setup, we examined bees’ preference between two flower types after having been trained …
Figure 1—figure supplement 3
Counterbalanced colour sets used in experiments 1–6.
Groups of bees were trained and tested with counterbalanced colour sets and training sequences in each experiment.
Figure 1—figure supplement 2
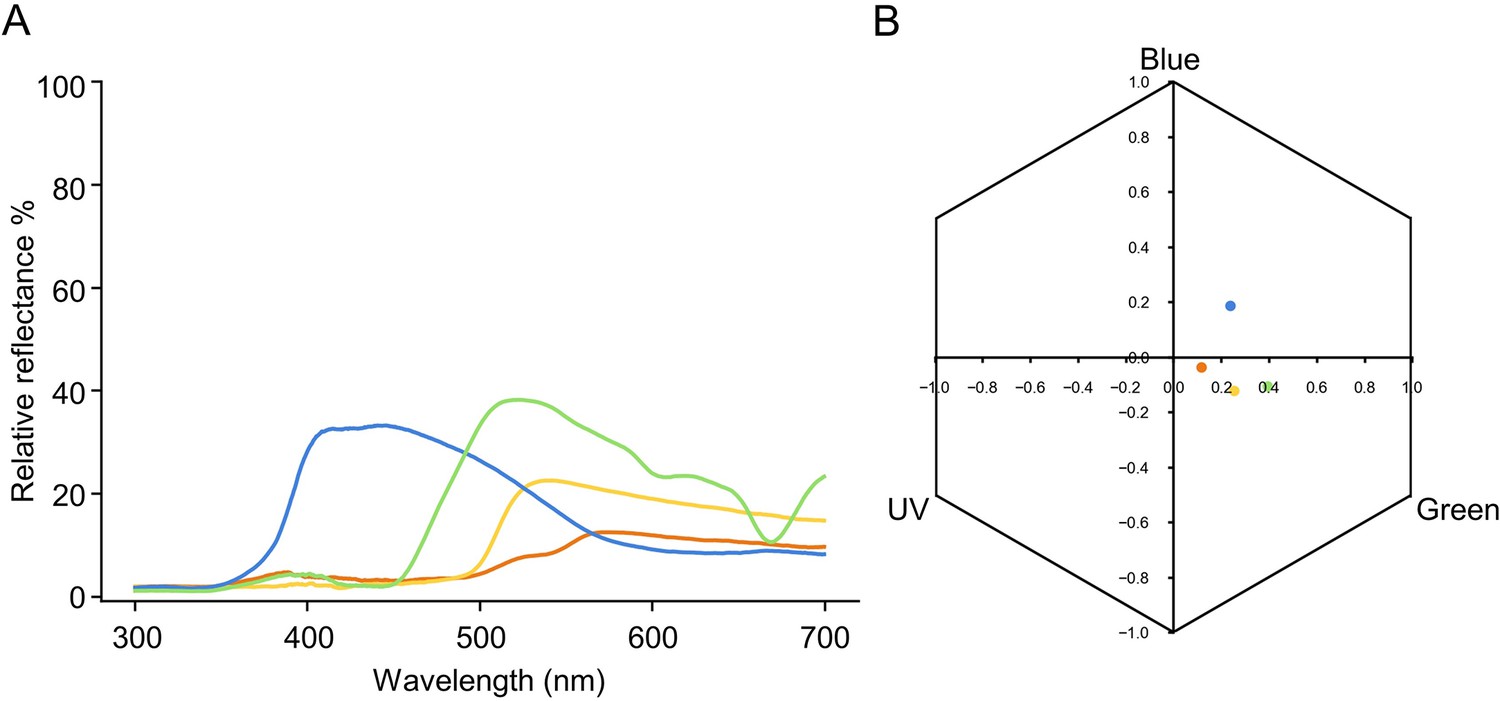
Specifications of the colours used for all the experiments.
(A) Spectral reflectance plot of blue, orange, yellow, and green colours used. (B) Loci of colours in the hexagonal bee colour space, determined by the responses each colour elicits on the bee’s UV, …
Figure 1—figure supplement 1
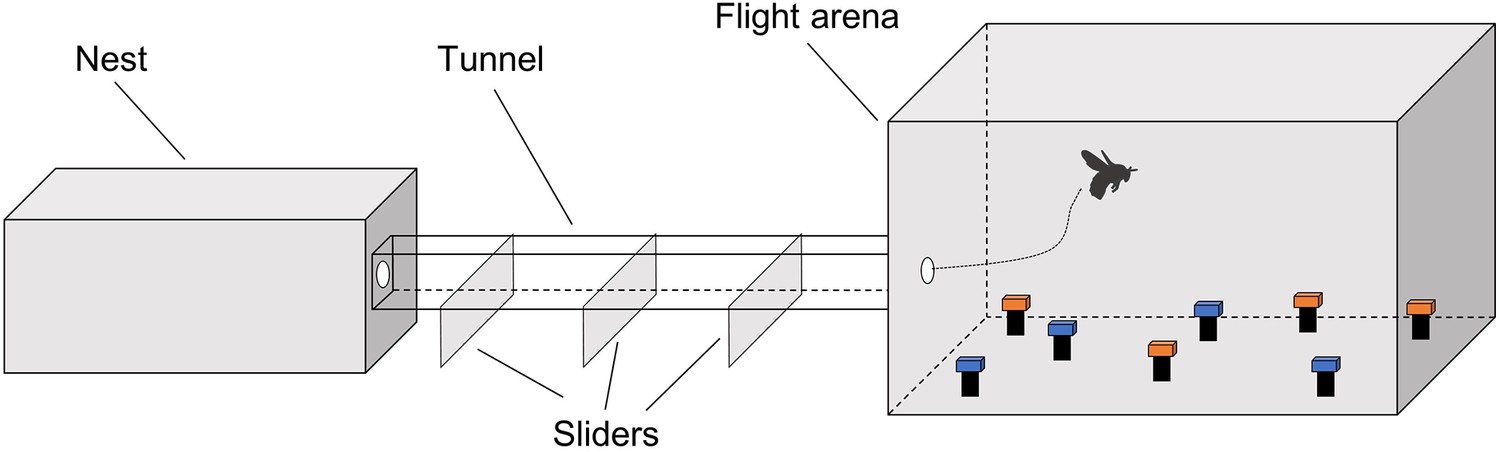
General setup for experiments 1, 2, and 3.
In each experiment, artificial flowers of paired colours were horizontally presented in the training and testing phase.
Results
If bumblebees encode and retrieve memories for absolute values, their preference for a particular option should not depend on the context in which that option was learned (Padoa-Schioppa and Assad, 2008). To test this idea, we conducted experiment 1 with a multi-contextual design where certain flowers had the same quality of reward but different ranking. Bees were first trained (individually in all experiments) on two different pairs of coloured flowers (A and B; C and D; Figure 1A, Figure 1—figure supplement 1, and Figure 1—figure supplement 2). The sucrose concentrations of the different pairs of flowers were chosen according to Weber’s Law (Akre and Johnsen, 2014) to represent the same perceived difference, i.e. A:B=45%:30% had the same contrast of incentives as C:D=30%:20% (Figure 1A). Note that for all experiments, the order of training sessions was counterbalanced to ensure that test preferences were not a result of a recency effect. Meanwhile, the focal colours used in the unrewarded test were counterbalanced during training to account for any colour effects (Materials and methods; Figure 1; Figure 1—figure supplement 3). Following sequential training in both contexts (Figure 1—figure supplement 3 and Figure 1—figure supplement 4), we tested each bee’s preference between flower types B and C, which had the same reward quality (30%) during training. Therefore, if bumblebees’ preferences were driven exclusively by their memory for absolute values for options, they should show no preference between B and C in the test. Any significant preference would suggest a contextual effect due to B and C having different rankings during training (B<A while C>D). In an unrewarding test with B and C flowers, bees significantly preferred C over B (generalised linear model [GLM]: 95% CI = [0.08, 0.38], N=40, and p=4.00e-3; Figure 1B and Figure 1—figure supplement 3). Although these results suggest that bees store memories for the relative ranking of flowers, they still may encode absolute information (Table 1), and therefore, we carried out additional experiments.
Table 1
Predictions for various decision strategies for bumblebees’ flower preferences in experiments 1, 2, and 3.
The left column lists the different categories of decision strategies. The middle three columns show the predicted results of each strategy for each experiment. The right column shows whether the …
| Strategy | Expected preference | Matches bees’ behaviour | ||||
|---|---|---|---|---|---|---|
| Experiment 1B (30%) vs C (30%) | Experiment 2B (30%) vs C (15%) | Experiment 3A (45%) vs C (30%) | ||||
| Absolute memory | Indifferent | B | A | ✗ | ||
| Remembered ranking | C | C | Indifferent | ✓ | ||
| Lexicographic combination—absolute memory priority | C | B | A | ✗ | ||
| Lexicographic combination—ranking priority | C | C | A | ✗ | ||
| Non-lexicographic combination | C | B/C/indifferent | A | ✗ | ||
| Value by association | B | B | A | ✗ | ||
| State-dependent valuation learning | C | C | C | ✗ | ||
Bees may still store and use memories for both absolute and relative information in some combined fashion. To test this, in experiment 2, we set the reward quality of flowers B and C to be different (Figure 1C and Figure 1—figure supplement 3). The sucrose concentration of flowers was A>B >> C>D (45%, 30%, 15%, and 10%). After training in both contexts, bees’ preference between B and C flowers was tested. If bumblebees retain and use memories for both ranking and absolute information (and the value weights for these two different types of information are not extremely unbalanced), bees should either prefer B over C because it is much higher in quality, or this value difference would cancel out the ranking difference (B<A while C>D), and their preference for B and C should be equal. On the other hand, a preference for the much lower reward quality flower C would indicate they have access to only ranking memories. Similar to experiment 1, the result of the unrewarding test showed that bees preferred C (GLM: 95% CI = [0.07, 0.28], N=40, and p=3.30e-3; Figure 1D).
Note that a preference for C over B in experiments 1 and 2 may have resulted from remembered ranking or a distorted absolute memory for flower type B due to the presence of and comparison with A during training. Evidence for the latter would manifest as a difference in the preference for C over B in experiment 2 compared to in experiment 1, whereas remembered ranking would predict a similar preference. There was no significant difference in preference for C across experiments 1 and 2 (GLM: 95% CI = [–0.24, 0.13], N=80, and p=0.55; Figure 1B and D). There is thus no evidence that bumblebees’ choices in novel contexts are based on remembered absolute metrics.
If bumblebees rely purely on remembered ranking, then their memories for options should only be ordinal (Vlaev et al., 2011). That is, they can tell one option is better than another but cannot tell how much better. If so, there should be no indication that bumblebees can compare options in quantitative terms. To test this, in experiment 3, we made the reward contrast ratios within the two contexts very different ([A:B] = [1.5:1]=45%:30% and [C:D] = [6:1]=30%:5%; A>B = C>>D; Figure 1E). Here, testing bees’ preference between A and C flowers can answer our question. If bees encode memories for relative differences in quantitative terms, C should be preferred over A, as C had a higher relative value than A. Alternatively, if bees only remember the ordinal rank of options, preference for A and C should be the same because they have the same rank within their training contexts. In the unrewarding test, bees chose A and C equally (GLM: 95% CI = [–0.16, 0.12], N=40, and p=0.79; Figure 1F), suggesting bumblebees store and recall only memories for the qualitative difference between options. Furthermore, bumblebees’ lack of preference for the equally ranked A and C flowers, despite their difference in sugar concentration, demonstrates absolute memories were not remembered (Table 1) and, in combination with experiments 1 and 2, suggests that bumblebees only encode and recall remembered ranking for use in novel contexts.
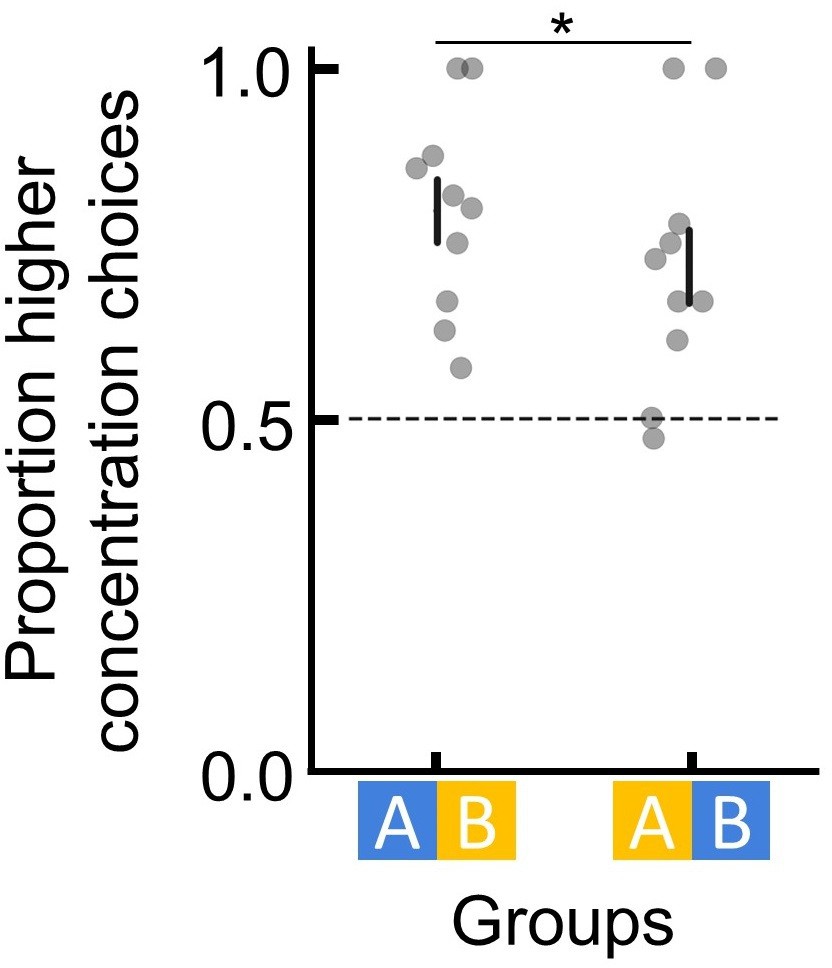
It may be that bumblebees can store and recall absolute information, but any ranking information available overshadows absolute memory. To test this, in experiment 4, we trained bees so that ranking information was absent. Bees experienced a temporally separated training procedure (Materials and methods; Figure 2A) whereby they first foraged on one flower with either 45% or 30% sugar solution, and 1 hr later they were trained on a different flower with either 30% or 45%, respectively. In the subsequent (1 hr later) unrewarded test, bees’ preferences were assessed. Because the two flower types were separated physically and temporally, bees had no ranking information available to putatively overshadow absolute information. Therefore, any preferences formed would be a result of remembered absolute information. However, bees had no significant preferences for either option (GLM: 95% CI = [–0.15, 0.22], N=20, and p=0.69; Figure 2B), suggesting that bees’ lack of absolute memory in the tests of experiments 1–3 was unlikely to be a result of absolute memories being less salient and overshadowed by ranking memories.
Figure 2
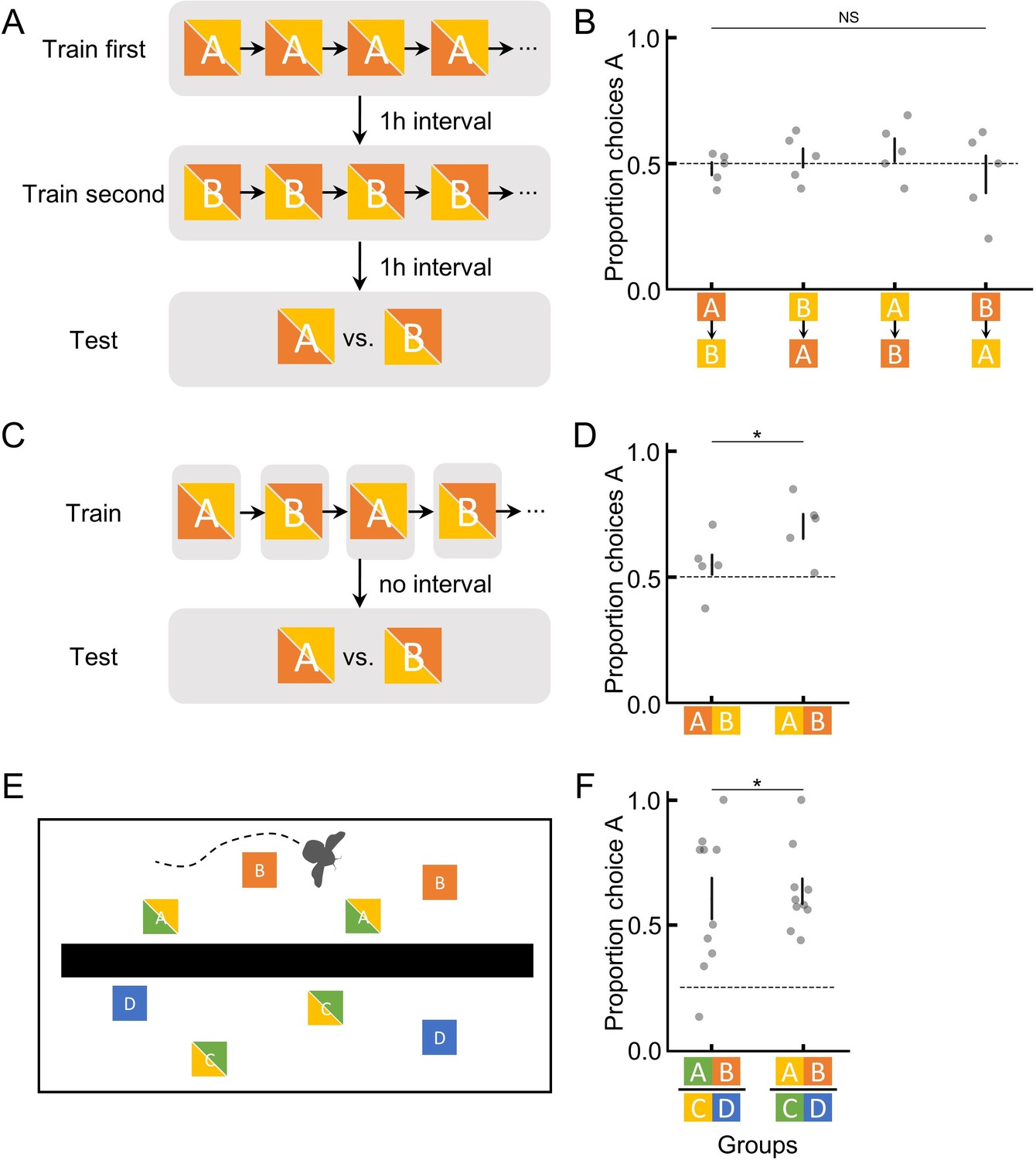
Bumblebees are only able to utilise absolute information for short periods of time in order to encode and recall ranking information in novel contexts.
(A) Illustration of the training and testing procedure in experiment 4, where two options were separately trained and tested, with a 1-hr interval in between to ensure clear spatial and temporal …
In order to discriminate any two sequentially visited flowers and indeed to rank them, bumblebees must utilise absolute information for at least a brief amount of time. During training in experiments 1–3, bumblebees could see flowers of both types simultaneously. Given the results of experiment 4, we therefore asked whether bumblebees could utilise absolute information long enough to rank flowers that were temporally close together but visually separated. To do this, in experiment 5, we trained bees on one flower for each bout, i.e. each visit to the arena (Materials and methods; Figure 2C). This way, both flower types were never in a bee’s visual field at the same time. During the unrewarded test, bees preferred the option that had offered the higher sugar concentration in training (GLM: 95% CI = [0.23, 0.87], N=10, and p=8.29e-3; Figure 2D), suggesting that absolute information about a flower learned in visual isolation can be retained for at least a couple of minutes but, as experiment 4 highlights, is eventually lost or not able to be recalled later. Experiment 4 and 5 thus collectively indicate that bumblebees can use absolute information to compare options only in their short-term memories (range of minutes) but not in their mid-term memories (range of hours; Menzel, 2001).
Our results highlight the potential disadvantage of using only remembered ranking, i.e. preferring to search for a less rewarding flower. How might our results speak to bumblebees’ behaviour in more ‘natural’ settings? In the wild, bumblebees often forage in situations whereby when drinking from one flower type they cannot see other flower types in their field of view, i.e. on the other side of a bush or many meters away. Yet, the distance between any two flowers within a bumblebee’s foraging range can be traversed within minutes (Osborne et al., 1999), i.e. within the range of bumblebees’ short-term memories (Menzel, 2001). Therefore, we would expect that bumblebees in the wild would not be susceptible to learning preferences for a less rewarding flower despite being unable to retain absolute information for very long. In an attempt to formally test this idea, in experiment 6, we trained bees in a multi-contextual semi-realistic foraging situation. Here, in a divided arena, bumblebees learned to forage from flowers of four different colours (Figure 2E and Figure 1—figure supplement 3) but could only see flowers of two colours at any one time, i.e. could only see A and B (45% and 30%) flowers from one side of the arena and could only see C and D (30% and 20%) flowers from the other side (Figure 2E). In the unrewarding test with all four options available, bees preferred option A (GLM: 95% CI = [0.94, 1.68], N=20, and p=1.21e-6; Figure 2F), indicating that the short retention/utilisation of absolute memories that bumblebees showed in our lab-created situations are likely not to be disadvantageous to bumblebees in the wild, given their natural behaviour and ecological niche.
Discussion
Our results suggest that bumblebees are only able to make use of ordinal ranking memories to guide foraging choices outside their original learning contexts. Although absolute information is essential for any animal to compare any two options initially, our findings show that only ordinal ranking can be recalled by bumblebees later in new contexts. Our results suggest that temporal adjacency is necessary for absolute information of different options to be compared by bees. That is, once bees experience one option, in order to utilise the absolute information, they must experience a second option within several minutes. Both our proof-of-concept experiment (experiment 6) and other previous multi-choice semi-realistic foraging tasks (e.g. Greggers and Menzel, 1993) show that with short inter-flower visits, bees can utilise absolute information in order to compare and rank flowers.
Another potential mechanism worth discussing which could potentially explain bumblebees’ preferences is state-dependent valuation learning (SDVL; for a review see McNamara et al., 2012; Table 1). If bees’ choices were a result of SDVL instead of remembered ordinal ranking, then their preferences should be quantitative, i.e. larger values should be assigned with larger differences between internal state and flower experience. However, the results of experiment 3 speak against this interpretation because bees were indifferent between equally ranked A and C even though they were experienced in a richer and poorer environment, respectively.
Why would bumblebees have evolved to use only memories for ordinal comparisons while humans and starlings evolved to retain and recall both absolute and ranking memories? Breadth of diet has been suggested to play a role in the evolution of cognition (Hemingway et al., 2017; MacLean et al., 2014; Simons and Tibbetts, 2019). Humans and starlings forage on a range of different foods, whereas adult bumblebees feed almost exclusively on nectar and pollen from flowers. Perhaps a varied diet, whereby one might need a common currency across vastly different food types (Chib et al., 2009; Levy and Glimcher, 2012) may have forced some animals to retain and use absolute memories, whereas a limited diet and limited dimensionality of reward might have favoured memories for ordinal relationships alone.
There is no reason yet to suggest a bee’s brain lacks the neural substrates to retain and recall absolute memories for options for later use in novel contexts. In fact, recent electrophysiological recordings on the gustatory neurons of bumblebees demonstrated that the spiking rates of these peripheral sensory neurons increased as a function of sucrose concentration (Miriyala et al., 2018). These gustatory neuronal signals could theoretically be used by higher centres of an animal’s brain to encode an option’s utility as a cardinal (absolute) value (Schultz, 2015). However, our results suggest that bumblebees do not access or utilise any potentially stored absolute memories when it would be of benefit in novel contexts. Given our results, it would be intriguing to determine the differences in the underlying neural mechanisms responsible for the utilisation of absolute properties and ordinal ranking memories. Whatever the ultimate and proximate causes of bumblebees’ ordinal-only memories for options’ values, our findings demonstrate a fundamental difference in the mechanisms underlying learned preferences between those of bumblebees and those of starlings and humans.
Materials and methods
Animals and setup
Request a detailed protocolBumblebee (Bombus terrestris) colonies were obtained from the Chinese branch of the Biobest Group (Biobest Belgium N.V., Westerlo, Belgium) and housed in wooden nest boxes (28 cm × 16 cm × 11 cm). Although there are no current requirements regarding insect care and use in research, experimental design and procedures were guided by the 3Rs principles (replacement, reduction, and refinement; Russell and Burch, 1959). The behavioural tests were non-invasive, and the types of manipulations used (sucrose and water) are all experienced by bumblebees during their natural foraging life in the wild. The bumblebees were cared for on a daily basis by trained and competent staff, which included routine monitoring of welfare and provision of correct and adequate food during the experimental period. A foraging arena (40 cm × 59 cm × 41 cm; Figure 1—figure supplement 1) was connected to the nest boxes via an acrylic tunnel with sliding doors, allowing experimenters to control bees’ access to the arena. Individual bees were marked with number tags (Opalithplättchen, Warnholz & Bienenvoigt, Ellerau, Germany), which were super-glued to the bees’ thorax. To ensure the sucrose solution concentration used in experiments was motivating, colonies were fed outside of experiments with 5–15% sucrose solution (w/w). They were also provided with ~3 g pollen every day. Illumination was provided by daylight fluorescent tubes (MASTER TL-D 90 DeLuxe 36 W/965, Philips, Eindhoven, the Netherlands) and near-UV fluorescent tubes (TL-D 36 W BLB, Philips) with high-frequency electronic ballasts (EB-Ci 1–2 36 W/1–4 18 W, 42–60 kHz, Philips) to generate a flicker frequency beyond the bumblebee’s flicker-fusion frequency. Coloured acrylic squares (25 mm × 5 mm × 5 mm) set on top of opaque glass cylinders as (artificial) flowers were placed in the arena with a different random spatial arrangement each trial. The spectral reflectance (Figure 1—figure supplement 2A) of all flower colours used in experiments was measured with a wavelength range of 300–700 nm and with 1 nm increments, using a spectrophotometer (Ocean Optics USB 2000+; Shanghai, China) and a deuterium/halogen light source. The perceptual positions of the colours in the bee colour hexagon space (Figure 1—figure supplement 2B) were calculated using the spectral reflectance measurements and the published Bombus terrestris spectral sensitivity functions of their three photoreceptors (Chittka, 1992; Skorupski et al., 2007). The minimal perceptual distance of all the colour combinations used in this study was above 0.14 hexagon units, which can be well differentiated by bumblebees (Dyer and Chittka, 2004). Furthermore, neurophysiological (Miriyala et al., 2018) and behavioural evidence (Whitney et al., 2008) verify that the sugar concentration differences used in this study can be readily discriminated by bumblebees.
Experimental protocol
All bees were individually pre-trained on eight transparent artificial flowers and were allowed to collect a full crop of sucrose solution from those flowers. Once a bee had successfully foraged for at least three consecutive bouts, she was moved to the training phase of an experiment.
Experiments 1, 2, and 3
Request a detailed protocolBees (N=40 for each experiment) were trained to forage from eight flowers, four of each of two colours (Figure 1—figure supplement 1 and Figure 1—figure supplement 3). In experiment 1, one group of bees (n=10) learned (individually) that green flowers contained 45% sucrose solution and that yellow flowers contained 30% sucrose solution. Once training on these two flower types was complete (more detail below), this group of bees learned (individually) that orange flowers contained 30% sucrose solution and that blue flowers contained 20% sucrose solution. Training for bees in experiments 2 and 3 was the same except the sucrose concentrations were different. Training sequences and colour combinations were equally counterbalanced across bees in each experiment (Figure 1—figure supplement 3). Flower colours and sequences for all experiments are listed in Figure 1—figure supplement 3. Initially during training, all bees would land and drink from both flowers. As bees learned the colour-reward contingency, they began to land on the less rewarded flowers but did not drink. Once this behaviour was observed, during subsequent bouts, we removed and replaced one higher-ranking flower with one lower-ranking flower. By doing so, and because none of the flowers were refilled during training bouts, bees would again visit the lower-ranking flowers and drink the sugar water droplet. This method helped us to ensure that bees experienced drinking sucrose solution from both options an equal number of times. Training was completed when a bee collected 50 of the 20 μl aliquots of sugar reward from flowers of both colours. Note that we verified, in our setup, that bees could learn to discriminate between flowers of different colours and sucrose concentrations (Figure 1—figure supplement 4). After training, each bee was individually tested with two types of flowers (four of each type) offering 20 μl droplets of unrewarding water, and all flower visits were recorded for 2 min. Note that in all tests, the two flowers types presented had not been experienced previously together in the same arena by the bees.
Experiment 4
Request a detailed protocolBees (N=20) were trained in the same way as the other experiments except that during training only one flower type was in the arena at any one time, i.e. either all of the flowers offered 45% sucrose solution or all flowers offered 30% sucrose solution (Figure 1—figure supplement 3; Figure 2A). Also, the inter-session interval for experiment 4 was 1 hr. Each session ended when the bee completed 50 flower visits. After training and after another 1 hr interval, bees underwent an unrewarded test with the two flower types they had been trained on, presented simultaneously in the arena. Both the colour/sucrose concentration contingency and the order of training sessions (which type of flower was trained last) were counterbalanced across bees (Figure 1—figure supplement 3; Figure 2B).
Experiment 5
Request a detailed protocolBees (n=10) were trained similarly to experiment 4 in which only one flower type was in the arena at any one time (Figure 1—figure supplement 3; Figure 2C). However, instead of two long sessions, bees experienced each flower in alternating bouts (visits to the arena to fill their crop). Once bees experienced 50 flower visits on each flower type, training ended and the bees underwent an unrewarded preference test with both flower types in the arena (Figure 1—figure supplement 3; Figure 2D).
Experiment 6
Request a detailed protocolBees (N=20) were trained and tested similarly to experiments 1–3 except that the two sets of flowers were in the arena at the same time and accessible to the bee during each bout. However, flowers were also separated by an opaque wall to prevent the bees from seeing both sets simultaneously (Figure 1—figure supplement 3; Figure 2E). The wall extended to cover most but not the full length of the arena, and there was space at the entrance and the far end of the arena so that bees could freely choose to visit flowers on either side of the wall. No matter where the bee was in the arena, she could not see both pairs of flowers at the same time. In addition, flowers were refilled after the bees emptied one and began drinking on another. Each group (n=10) was trained with either the yellow or green flower as the high concentration flower. The training was deemed complete when either a bee performed 200 landings or when a bee only landed exclusively on any one option for three consecutive bouts (note that bees never landed exclusively on the lower rewarding option for three bouts consecutively). After training, the bees were individually tested with all four flowers (two of each type) presented in the same arena with the opaque barrier removed (Figure 1—figure supplement 3; Figure 2F).
Statistical analyses
Request a detailed protocolR v.3.6.1 was used to perform all GLMs with quasibinomial distribution and logit link function. For all models, the response variable was the proportion of choices for the flower that had been associated with high concentration sugar water. Fixed factors for models for results of experiments were (i) colony of each bee, (ii) flower colour, and (iii) training sequence (whether the higher concentration sugar reward was used during the first or second training session). Significance of fixed effects was tested using likelihood ratio tests, and none were found to have a significant effect on bees’ preferences during the tests in any experiment. When comparing preference for flower C across experiments 1 and 2, experiment was set as a fixed factor.
Data availability
All data generated or analysed during this study are available on the Dryad Digital Repository: https://doi.org/10.5061/dryad.rr4xgxdb9.
-
Dryad Digital RepositoryBumblebees retrieve only the ordinal ranking of foraging options when comparing memories obtained in distinct settings.https://doi.org/10.5061/dryad.rr4xgxdb9
References
-
Psychophysics and the evolution of behaviorTrends in Ecology & Evolution 29:291–300.https://doi.org/10.1016/j.tree.2014.03.007
-
Context-dependent foraging choices in risk-sensitive starlingsAnimal Behaviour 64:251–260.https://doi.org/10.1006/anbe.2002.3059
-
Rigorously testing multialternative decision field theory against random utility modelsJournal of Experimental Psychology. General 143:1331–1348.https://doi.org/10.1037/a0035159
-
Evidence for a common representation of decision values for dissimilar goods in human ventromedial prefrontal cortexThe Journal of Neuroscience 29:12315–12320.https://doi.org/10.1523/JNEUROSCI.2575-09.2009
-
The colour hexagon: a chromaticity diagram based on photoreceptor excitations as a generalized representation of colour opponencyJournal of Comparative Physiology A 170:533–543.https://doi.org/10.1007/BF00199331
-
Biological significance of distinguishing between similar colours in spectrally variable illumination: bumblebees (Bombus terrestris) as a case studyJournal of Comparative Physiology. A, Neuroethology, Sensory, Neural, and Behavioral Physiology 190:105–114.https://doi.org/10.1007/s00359-003-0475-2
-
Memory dynamics and foraging strategies of honeybeesBehavioral Ecology and Sociobiology 32:17–29.https://doi.org/10.1007/BF00172219
-
Transitive or not: A critical appraisal of transitive inference in animalsEthology : Formerly Zeitschrift Fur Tierpsychologie 119:703–726.https://doi.org/10.1111/eth.12124
-
Rationality in decision-making in the fringe-lipped bat, trachops cirrhosusBehavioral Ecology and Sociobiology 71:94.https://doi.org/10.1007/s00265-017-2321-5
-
Context-dependent preferences in wild fruit batsAnimal Behaviour 179:65–72.https://doi.org/10.1016/j.anbehav.2021.06.016
-
Context-sensitive valuation and learningCurrent Opinion in Behavioral Sciences 41:122–127.https://doi.org/10.1016/j.cobeha.2021.05.001
-
The root of all value: a neural common currency for choiceCurrent Opinion in Neurobiology 22:1027–1038.https://doi.org/10.1016/j.conb.2012.06.001
-
The ecological rationality of state-dependent valuationPsychological Review 119:114–119.https://doi.org/10.1037/a0025958
-
Searching for the memory trace in a mini-brain, the honeybeeLearning & Memory 8:53–62.https://doi.org/10.1101/lm.38801
-
Burst firing in bee gustatory neurons prevents adaptationCurrent Biology 28:1585–1594.https://doi.org/10.1016/j.cub.2018.03.070
-
Context-dependent decisions among options varying in a single dimensionBehavioural Processes 89:115–120.https://doi.org/10.1016/j.beproc.2011.08.017
-
A landscape-scale study of bumble bee foraging range and constancy, using harmonic radarJournal of Applied Ecology 36:519–533.https://doi.org/10.1046/j.1365-2664.1999.00428.x
-
Context-dependent outcome encoding in human reinforcement learningCurrent Opinion in Behavioral Sciences 41:144–151.https://doi.org/10.1016/j.cobeha.2021.06.006
-
Rhesus macaques (Macaca mulatta) exhibit the decoy effect in a perceptual discrimination taskAttention, Perception & Psychophysics 77:1715–1725.https://doi.org/10.3758/s13414-015-0885-6
-
Paradox, performance, and the architecture of decision-making in animalsAmerican Zoologist 36:518–529.https://doi.org/10.1093/icb/36.4.518
-
Neuronal reward and decision signals: from theories to dataPhysiological Reviews 95:853–951.https://doi.org/10.1152/physrev.00023.2014
-
Context-dependent violations of rational choice in honeybees (Apis mellifera) and gray jays (Perisoreus canadensis)Behavioral Ecology and Sociobiology 51:180–187.https://doi.org/10.1007/s00265-001-0420-8
-
Insects as models for studying the evolution of animal cognitionCurrent Opinion in Insect Science 34:117–122.https://doi.org/10.1016/j.cois.2019.05.009
-
Photoreceptor spectral sensitivity in island and mainland populations of the bumblebee, Bombus terrestrisJournal of Comparative Physiology. A, Neuroethology, Sensory, Neural, and Behavioral Physiology 193:485–494.https://doi.org/10.1007/s00359-006-0206-6
-
The elusiveness of context effects in decision makingTrends in Cognitive Sciences 25:843–854.https://doi.org/10.1016/j.tics.2021.07.011
-
Not just for consumers: context effects are fundamental to decision makingPsychological Science 24:901–908.https://doi.org/10.1177/0956797612464241
-
Transitive inference in non-human animals: an empirical and theoretical analysisBehavioural Processes 78:313–334.https://doi.org/10.1016/j.beproc.2008.02.017
-
Does the brain calculate value?Trends in Cognitive Sciences 15:546–554.https://doi.org/10.1016/j.tics.2011.09.008
Decision letter
-
Giorgio VallortigaraReviewing Editor; University of Trento, Italy
-
Christian RutzSenior Editor; University of St Andrews, United Kingdom
-
Alex KacelnikReviewer; University of Oxford, United Kingdom
Our editorial process produces two outputs: (i) public reviews designed to be posted alongside the preprint for the benefit of readers; (ii) feedback on the manuscript for the authors, including requests for revisions, shown below. We also include an acceptance summary that explains what the editors found interesting or important about the work.
Decision letter after peer review:
Thank you for submitting your article "Bumblebees retrieve only the ordinal ranking of foraging options when comparing memories obtained in distinct settings" for consideration by eLife.
Your article has been reviewed by three peer reviewers, one of whom is a member of our Board of Reviewing Editors, and the evaluation has been overseen by Christian Rutz as the Senior Editor. The following individual involved in the review of your submission has agreed to reveal their identity: Alex Kacelnik (Reviewer #3).
The reviewers have discussed their reviews with one another, and the Guest Reviewing Editor has drafted this decision letter to help you prepare a revised submission.
Essential revisions:
While all three reviewers found merit in your paper, they raised several important concerns, indicating a need for conducting additional experiments. Here is a summary of the main issues:
Reviewer #1 suggests that the results of the first two experiments could be accounted for by a recency effect, where the higher item of the last comparison is better retrieved, and that in the last experiment, bumblebees' choice for the higher-ranking item could be based on its higher absolute quantitative value in terms of sucrose solution.
Reviewer #2 asks for an experimental assessment of whether bees do respond differently to the different colors and concentrations used. Moreover, Reviewer #2, as well as Reviewer #1, noted that the huge literature on transitive inference in non-human animals could be relevant to the topic addressed by the authors and deserves to be considered.
Reviewer #3, similarly to Reviewer #1, is not convinced that the authors can exclude sensitivity to absolute properties, and suggests some important control experiments, the most important one being the following: training of bees with only A and only B, each in a context, and then giving a choice: if bees show a preference when the only available information is concentration, then the argument that they just cannot remember concentrations could be rejected.
Several other, more specific, comments were provided by the reviewers and should be taken into account by the authors in a revision. In conclusion, the reviewers feel that the paper is potentially publishable but more experimental work is needed to address several issues raised by these fascinating experiments.
Note: Please note that eLife has adopted the STRANGE framework, to help improve reporting standards and reproducibility in animal behaviour research. In your revision, please consider scope for sampling biases and potential limitations to the generalisability of your findings:
https://reviewer.elifesciences.org/author-guide/journal-policies
https://doi.org/10.1038/d41586-020-01751-5
Reviewer #1 (Recommendations for the authors):
It seems to me that the study is interesting and highlights some fascinating ability of bees to adapt and implement different strategic behaviour to better succeed in different tasks.
Reviewer #2 (Recommendations for the authors):
Line 61-63: "Only recently" refers to the work of Pompilio and Kacelnik (1), which is already 12 years old. This does not seem "recent".
Line 63: Please avoid superlative qualifications ("impressive" studies).
Line 70: The name REAL in capitals should be changed to the appropriate citation format.
Lines 118-124: The explanations provided for the lexicographic and the non-lexicographic combinations strategies are not intuitive and difficult to follow both for advised and unadvised readers. Please, be more didactic in your explanations.
Line 200-201: In fact, the work of Greggers and Menzel (1) shows that honey bees can memorize simultaneously four different feeding options providing different reward qualities (sucrose flow rate) and labeled with different colors (and they match their choice to their feeding properties). Mentioning/discussing this would be useful.
Line 205: I could not understand why you are citing here the work of Miriyala et al., (2), which is a work on electrophysiological properties of gustatory receptors with no relation to brain encoding of utility. Miriyala et al., measured peripheral gustatory activity, not central encoding of gustatory information.
References
1. U. Greggers, R. Menzel, Memory dynamics and foraging strategies of honeybees. Behavioral Ecology and Sociobiology 32, 17-29 (1993).
2. A. Miriyala, S. Kessler, F. C. Rind, G. A. Wright, Burst firing in bee gustatory neurons prevents adaptation. Curr Biol 28, 1585-1594 (2018).
Reviewer #3 (Recommendations for the authors):
L87: "if bumblebees remembered absolute values for options, their preference between B and C in the test should be identical"
The word 'remembered' should be changed to 'were driven exclusively by their memory for'. An animal can 'remember' something but not use only this memory when other cues are available, as it appears elsewhere in the paper.
L97, Figure 1: It would have been nice for completeness to have a control where bees were trained with A(45) and C(30) in the absence of any competitor flower types, and then offered a choice of A vs. C. I assume that they would prefer A, but if the authors are correct in claiming that bees don't retain absolute memories, even in that case the bees would be indifferent. If the bees in such a control preferred A, this would show that they DO remember quality when other cues are not competing. As it is, the data are compatible with attention to absolute concentration being overshadowed by information about ranking, rather than bees being unable to remember absolute values.
L130: "If bumblebees have memories for both ranking and absolute information, they should either prefer B over C because it is much higher in quality, or this value difference would cancel out the ranking difference (B < A while C > D) and their preference for B and C should be equal. On the other hand, a preference for the much lower reward quality flower C would indicate they have access to only ranking information."
This logic is not completely tight. If, say, the difference in preference induced by absolute memories were X, and that induced by ranking were Y, where X <
L142: "remembered ranking would predict a similar preference. There was no difference in preference for C across experiments 1 and 2 (GLM: 95% CI = [-0.24 0.13], N = 80, P = 0.55; Figure 1B and D)."
From visual inspection of Figure 1B and D it appears that there is in fact a small (and noisy) greater preference in 1B than in 1C. The analysis presented shows that this is not statistically significant. The authors interpret that lack of significance as meaning lack of a difference. This should be tempered, for the usual reasons: lack of significance is not enough to confirm that the null hypothesis (lack of effect) is actually true.
L162 "Further, bumblebees' equal preference for the equally ranked A and C flowers, despite their difference in sugar concentration, demonstrates absolute memories were not used (Table 1) and, in combination with experiments 1 and 2, confirms that bumblebees only use remembered ranking."
I am not convinced. Say that an animal encounters flowers A and B in a random sequence. It then assigns a value to each according to their ranking in that context (A > B), regardless of their absolute concentration. But, each time the animal encounters B, how does it know that it is worse than A? It must be because it DOES remember A, otherwise it could not establish a ranking. In other words, bees may not use, or may not remember, absolute values when moving to new contexts, but they must be sensitive to absolute values, at least within a context. So, maybe there is only short-term memory for absolute value, or maybe that is erased (reset) whenever a new context is entered, but the task of ranking is impossible without some memory for concentration existing and being used.
Again, the simple control of training with only A and only B, each in a context, and then giving a choice would be highly informative: if they show a preference when the only available information is concentration, then the argument that bumblebees just cannot remember concentrations could be rejected.
L182 "In the unrewarding test with all four options available, bees preferred option A (GLM: 95% CI = [0.94,1.68], N = 20, P = 1.21e-06; Figure 2B), indicating that despite being susceptible to suboptimal outcomes in lab-created situations, using only memories of options' ordinal ranking may be evolutionarily rational, i.e. can still lead to optimal choices within an ecologically relevant scenario."
I don't think the language here is used tightly. What Exp 4 shows is that while in 2-way choices bees showed strong context effects (Figure 1B), in 4-way choices animals' preferences match the use of absolute memory as well as ranking. Exp 4 is presented as if the only difference with the earlier design were the training condition, but it also differs in the choice procedure (4 vs. 2 options). It is known that other animals are sensitive to the number of options at choice time. The claim that the situation in Exp 4 is more ecologically relevant than that in the previous experiments is not fully explained, and it would be good to make it more convincing. As it is, it appears somewhat post-hoc.
As elsewhere in the paper, the use of the words optimal and sub-optimal is a bit loose. Any partial preference is suboptimal, because an 'optimal' animal (which is a theoretical construct) allocates all its behaviour to the best option in the context. In this article, following a practice in many experimental psychology writings, the words are used to mean any bias in the favourable direction, regardless of its magnitude. How would one call a theoretical bee that showed exclusive preference for the best option, 'super-optimal', perhaps? By definition, it should not be possible to better optimal behaviour. One could simply say that the psychology of the animals may be adaptive under natural conditions, not 'optimal'. In truth, I even don't like this, because we can't test whether animals are well designed or not. Optimality studies test models, not the notion that animals are well adapted.
L279: "The training was deemed complete when either a bee performed 200 landings or when a bee only landed on the mostly rewarded option for three consecutive bouts."
This method could produce an artefactual bias. If a bee did 3 bouts in a flower other than the 'most rewarded', then the test continues, but if it does it in the richest one, the test stops. This traps good results that could occur at random in any sequence. Under the null hypothesis that bees have no preference, this would show a bias towards the most rewarded flower because the data are censored when a good response is shown. It is equivalent to keep adding individuals to a sample until a significant result is observed, and then stop. I assume that bees may never have shown 3 bouts on a lesser flower, and if this is the case, please state so.
https://doi.org/10.7554/eLife.78525.sa1Author response
Essential revisions:
While all three reviewers found merit in your paper, they raised several important concerns, indicating a need for conducting additional experiments. Here is a summary of the main issues:
Reviewer #1 suggests that the results of the first two experiments could be accounted for by a recency effect, where the higher item of the last comparison is better retrieved, and that in the last experiment, bumblebees' choice for the higher-ranking item could be based on its higher absolute quantitative value in terms of sucrose solution.
In the first three experiments, the order in which the pairs of different sucrose concentrations were used across training sessions was counterbalanced (listed in Figure 1— figure supplement 3). Further, the results of our GLMs show that the order of training had no effect on preferences. Therefore, the results of the first two experiments cannot be explained by a recency effect. We have highlighted these points on lines 101 – 105.
As we alluded to in our original submission, and as mentioned by Reviewer #3, bees must be able to assess the absolute properties for at least some short amount of time, otherwise they could not even discriminate sequentially visited flowers. Our work shows that bumblebees either do not retain memory for (or do not utilise encoded) absolute information for very long. We have now made these points clearer in our revised manuscript.
With the addition of new data from experiments which address Reviewer #3’s concerns, we have also provided a clearer rationale and interpretation of experiment 6 (last experiment in our original submission). Please see our responses to Reviewers below for more detail.
Reviewer #2 asks for an experimental assessment of whether bees do respond differently to the different colors and concentrations used.
In our revised manuscript, we now provide support through previous works and our own experiments (original and newly added) that bumblebees can easily discriminate between the colours used and between the sugar concentrations used in our experiments on lines 348 – 353. Please see our response to Reviewer #2’s comment #1 for more detail.
Moreover, Reviewer #2, as well as Reviewer #1, noted that the huge literature on transitive inference in non-human animals could be relevant to the topic addressed by the authors and deserves to be considered.
Transitive inference (TI) is only superficially related to our question and methods applied here. In short, TI tests for the ability to infer relative ranking and requires subjects not to learn absolute information. We tested whether and to what extent bumblebees remember absolute and/or relative ranking information. Nonetheless, we agree that it may be useful to mention briefly how and why these questions and designs are different. Therefore, we have now included a paragraph for this purpose in the Introduction on lines 73 – 89. Please also see our response to Reviewer #2's comment #3 below for more details.
Reviewer #3, similarly to Reviewer #1, is not convinced that the authors can exclude sensitivity to absolute properties, and suggests some important control experiments, the most important one being the following: training of bees with only A and only B, each in a context, and then giving a choice: if bees show a preference when the only available information is concentration, then the argument that they just cannot remember concentrations could be rejected.
This was a great suggestion. We have done the experiment and have shown that bees form no preference when ranking information was unavailable. We discuss the details of this experiment in our response to Reviewer #3 below and in the revised manuscript on lines 204 – 233, and 385 – 401.
Several other, more specific, comments were provided by the reviewers and should be taken into account by the authors in a revision. In conclusion, the reviewers feel that the paper is potentially publishable but more experimental work is needed to address several issues raised by these fascinating experiments.
We have also addressed all specific comments below and hope that you and all reviewers find the revised manuscript acceptable for publication.
Note: Please note that eLife has adopted the STRANGE framework, to help improve reporting standards and reproducibility in animal behaviour research. In your revision, please consider scope for sampling biases and potential limitations to the generalisability of your findings:
https://reviewer.elifesciences.org/author-guide/journal-policies
https://doi.org/10.1038/d41586-020-01751-5
Reviewer #1 (Recommendations for the authors):
It seems to me that the study is interesting and highlights some fascinating ability of bees to adapt and implement different strategic behaviour to better succeed in different tasks.
Thank you!
Reviewer #2 (Recommendations for the authors):
Line 61-63: "Only recently" refers to the work of Pompilio and Kacelnik (1), which is already 12 years old. This does not seem "recent".
Fair point. We wanted to contrast these works (Pompilio and Kacelnik 2010 as well as Bavard et al., 2021 and 2018, and Klein et al., 2017) with those looking at contextual effects, which began 20 and more years ago. To better reflect our intended meaning, we therefore now state “Only more recently”.
Line 63: Please avoid superlative qualifications ("impressive" studies).
Yes, agreed, deleted.
Line 70: The name REAL in capitals should be changed to the appropriate citation format.
Fixed.
Lines 118-124: The explanations provided for the lexicographic and the non-lexicographic combinations strategies are not intuitive and difficult to follow both for advised and unadvised readers. Please, be more didactic in your explanations.
We have now provided a clearer explanation for all the strategies on lines 143 – 162.
Line 200-201: In fact, the work of Greggers and Menzel (15) shows that honey bees can memorize simultaneously four different feeding options providing different reward qualities (sucrose flow rate) and labeled with different colors (and they match their choice to their feeding properties). Mentioning/discussing this would be useful.
We have now included reference of this study in the Discussion on lines 279 – 282. Note that the Greggers and Menzel study, in which four feeding stations were simultaneously present in close proximity to each other, was not designed to determine whether bees remember absolute versus ranking information. But it is similar to our experiment 6 in that it provides a multicontextual semi-realistic foraging situation where the flowers can be visited quickly but cannot be seen together all at once.
Line 205: I could not understand why you are citing here the work of Miriyala et al., (16), which is a work on electrophysiological properties of gustatory receptors with no relation to brain encoding of utility. Miriyala et al., measured peripheral gustatory activity, not central encoding of gustatory information.
We apologise for the confusion, and thank you for highlighting this. We meant to cite Miriyala et al., to point out that information on fine sugar concentration differences are encoded in the peripheral gustatory neural responses. We now have made this clear on lines 302 – 305.
Reviewer #3 (Recommendations for the authors):
L87: "if bumblebees remembered absolute values for options, their preference between B and C in the test should be identical"
The word 'remembered' should be changed to 'were driven exclusively by their memory for'. An animal can 'remember' something but not use only this memory when other cues are available, as it appears elsewhere in the paper.
True, thanks. Changed.
L97, Figure 1: It would have been nice for completeness to have a control where bees were trained with A(45) and C(30) in the absence of any competitor flower types, and then offered a choice of A vs. C. I assume that they would prefer A, but if the authors are correct in claiming that bees don't retain absolute memories, even in that case the bees would be indifferent. If the bees in such a control preferred A, this would show that they DO remember quality when other cues are not competing. As it is, the data are compatible with attention to absolute concentration being overshadowed by information about ranking, rather than bees being unable to remember absolute values.
Thank you again for this suggestion! As mentioned above in response to your other comments, our new experiments show that when ranking information is absent, i.e. bees were trained on one flower at a time, bees were unable to retain or utilise memories for absolute information beyond the range of their short-term memories (for a few minutes but much less than one hour). Together, now with the help of the new experiments 4 and 5, our results show that bumblebees only retain ordinal ranking information in the long term and for use in novel contexts, and unless the temporal separation between flower experiences is relatively short, bumblebees are unable to use absolute information to even rank flowers. We include description and discussion of the additional experiments 4 and 5 on lines 204 – 233, 385 – 401, and in Figure 2.
L130: "If bumblebees have memories for both ranking and absolute information, they should either prefer B over C because it is much higher in quality, or this value difference would cancel out the ranking difference (B < A while C > D) and their preference for B and C should be equal. On the other hand, a preference for the much lower reward quality flower C would indicate they have access to only ranking information."
This logic is not completely tight. If, say, the difference in preference induced by absolute memories were X, and that induced by ranking were Y, where X <
We agree the logic was not totally solid, therefore we have changed the first sentence to the following (on lines 168 – 173):
“If bumblebees retain and use memories for both ranking and absolute information (and the value weights for these two different types of information are not extremely unbalanced), bees should either prefer B over C because it is much higher in quality, or this value difference would cancel out the ranking difference (B < A while C > D) and their preference for B and C should be equal.”
L142: "remembered ranking would predict a similar preference. There was no difference in preference for C across experiments 1 and 2 (GLM: 95% CI = [-0.24 0.13], N = 80, P = 0.55; Figure 1B and D)."
From visual inspection of Figure 1B and D it appears that there is in fact a small (and noisy) greater preference in 1B than in 1C. The analysis presented shows that this is not statistically significant. The authors interpret that lack of significance as meaning lack of a difference. This should be tempered, for the usual reasons: lack of significance is not enough to confirm that the null hypothesis (lack of effect) is actually true.
Fair point. We have now tempered this section by changing the wording on lines (181 – 185).
L162 "Further, bumblebees' equal preference for the equally ranked A and C flowers, despite their difference in sugar concentration, demonstrates absolute memories were not used (Table 1) and, in combination with experiments 1 and 2, confirms that bumblebees only use remembered ranking."
I am not convinced. Say that an animal encounters flowers A and B in a random sequence. It then assigns a value to each according to their ranking in that context (A > B), regardless of their absolute concentration. But, each time the animal encounters B, how does it know that it is worse than A? It must be because it DOES remember A, otherwise it could not establish a ranking. In other words, bees may not use, or may not remember, absolute values when moving to new contexts, but they must be sensitive to absolute values, at least within a context. So, maybe there is only short-term memory for absolute value, or maybe that is erased (reset) whenever a new context is entered, but the task of ranking is impossible without some memory for concentration existing and being used.
Again, the simple control of training with only A and only B, each in a context, and then giving a choice would be highly informative: if they show a preference when the only available information is concentration, then the argument that bumblebees just cannot remember concentrations could be rejected.
We apologise for the confusion, as we were not trying to claim that bumblebees are completely insensitive to absolute information. As you rightly point out, this would make comparing sequentially visited flowers with different sugar concentrations impossible to compare. We now point this out explicitly in our revised manuscript, both in the Abstract and the Results.
Thank you again for the suggested experiment. As we replied above, we now provide data from additional experiments showing that when learning only one flower at a time, bees only retain (or utilise memory of) absolute information for a short amount of time (at least a few minutes but much less than an hour). We now include a description of these experiments on lines 204 – 233, 385 – 401 and in Figure 2. L182 "In the unrewarding test with all four options available, bees preferred option A (GLM: 95% CI = [0.94,1.68], N = 20, P = 1.21e-06; Figure 2B), indicating that despite being susceptible to suboptimal outcomes in lab-created situations, using only memories of options' ordinal ranking may be evolutionarily rational, i.e. can still lead to optimal choices within an ecologically relevant scenario."
I don't think the language here is used tightly. What Exp 4 shows is that while in 2-way choices bees showed strong context effects (Figure 1B), in 4-way choices animals' preferences match the use of absolute memory as well as ranking. Exp 4 is presented as if the only difference with the earlier design were the training condition, but it also differs in the choice procedure (4 vs. 2 options). It is known that other animals are sensitive to the number of options at choice time. The claim that the situation in Exp 4 is more ecologically relevant than that in the previous experiments is not fully explained, and it would be good to make it more convincing. As it is, it appears somewhat post-hoc.
We apologise for the lack of explanation. Our intent was to simply determine whether the presumed strategy taken from the results of experiments 1-3 would lead to bees preferring flowers with lower sugar concentrations when presented in a situation more like they would experience in the wild. Bees forage in situations where different flowers can be seen in their visual field at the same time, but they can and will visit flowers which cannot be seen together with the other visted flowers (i.e. on the opposite side of a bush, or separated by many meters). Essentially, the design of our experiment 6 (previously experiment 4) mimics wild situations where bees drink from various flowers in different spatial locations whereby they cannot see all the different flowers at the same time but the duration between visits to different flowers is short (several seconds to a few minutes). We now explain this in more detail on lines 249 – 261.
With regards to the interpretation of experiment 6 (previously experiment 4), we believe that the addition of the new data from our experiments which address your comments above (now experiments 4 and 5) clearly indicate that bees’ preference differences between experiments 1-3 compared to experiment 6 are due to temporal differences between flower visits. As you rightly mentioned in your other comments, we now highlight more clearly in the revised manuscript that bumblebees must use absolute information to compare, and rank, any two sequentially visited flowers. However, our results suggest that bumblebees either do not retain or utilise absolute information beyond the range of their short-term memories. We now address this more clearly and with additional data on lines 204 – 233.
As elsewhere in the paper, the use of the words optimal and sub-optimal is a bit loose. Any partial preference is suboptimal, because an 'optimal' animal (which is a theoretical construct) allocates all its behaviour to the best option in the context. In this article, following a practice in many experimental psychology writings, the words are used to mean any bias in the favourable direction, regardless of its magnitude. How would one call a theoretical bee that showed exclusive preference for the best option, 'super-optimal', perhaps? By definition, it should not be possible to better optimal behaviour. One could simply say that the psychology of the animals may be adaptive under natural conditions, not 'optimal'. In truth, I even don't like this, because we can't test whether animals are well designed or not. Optimality studies test models, not the notion that animals are well adapted.
We agree and have now removed the terms optimality/suboptimality. We now use, where appropriate, advantageous/disadvantageous.
L279: "The training was deemed complete when either a bee performed 200 landings or when a bee only landed on the mostly rewarded option for three consecutive bouts."
This method could produce an artefactual bias. If a bee did 3 bouts in a flower other than the 'most rewarded', then the test continues, but if it does it in the richest one, the test stops. This traps good results that could occur at random in any sequence. Under the null hypothesis that bees have no preference, this would show a bias towards the most rewarded flower because the data are censored when a good response is shown. It is equivalent to keep adding individuals to a sample until a significant result is observed, and then stop. I assume that bees may never have shown 3 bouts on a lesser flower, and if this is the case, please state so.
Bees never went three bouts or more on the more poorly rewarding flower. We now state this in the Methods section on lines 413 – 415.
https://doi.org/10.7554/eLife.78525.sa2Article and author information
Author details
Funding
National Natural Science Foundation of China (31700988)
- Fei Peng
National Natural Science Foundation of China (31970994)
- Fei Peng
Key-Area Research and Development Program of Guangdong Province, China (2018B030340001)
- Fei Peng
Templeton World Charity Foundation (TWCF-2020-0539)
- Andrew B Barron
China Scholarship Council (202008440515)
- Yonghe Zhou
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Senior Editor
- Christian Rutz, University of St Andrews, United Kingdom
Reviewing Editor
- Giorgio Vallortigara, University of Trento, Italy
Reviewer
- Alex Kacelnik, University of Oxford, United Kingdom
Publication history
- Received: March 10, 2022
- Preprint posted: April 7, 2022 (view preprint)
- Accepted: September 12, 2022
- Version of Record published: September 27, 2022 (version 1)
Copyright
© 2022, Solvi, Zhou et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 999
- Page views
-
- 189
- Downloads
-
- 1
- Citations
Article citation count generated by polling the highest count across the following sources: PubMed Central, Crossref, Scopus.
Download links
Further reading
-
- Ecology
- Epidemiology and Global Health
The global spread of antibiotic resistance could be due to a number of factors, and not just the overuse of antibiotics in agriculture and medicine as previously thought.
-
- Ecology
- Evolutionary Biology
How the ecological process of community assembly interacts with intra-species diversity and evolutionary change is a longstanding question. Two contrasting hypotheses have been proposed: Diversity Begets Diversity (DBD), in which taxa tend to become more diverse in already diverse communities, and Ecological Controls (EC), in which higher community diversity impedes diversification. Previously, using 16S rRNA gene amplicon data across a range of microbiomes, we showed a generally positive relationship between taxa diversity and community diversity at higher taxonomic levels, consistent with the predictions of DBD (Madi et al., 2020). However, this positive 'diversity slope' plateaus at high levels of community diversity. Here we show that this general pattern holds at much finer genetic resolution, by analyzing intra-species strain and nucleotide variation in static and temporally sampled metagenomes from the human gut microbiome. Consistent with DBD, both intra-species polymorphism and strain number were positively correlated with community Shannon diversity. Shannon diversity is also predictive of increases in polymorphism over time scales up to ~4-6 months, after which the diversity slope flattens and becomes negative – consistent with DBD eventually giving way to EC. Finally, we show that higher community diversity predicts gene loss at a future time point. This observation is broadly consistent with the Black Queen Hypothesis, which posits that genes with functions provided by the community are less likely to be retained in a focal species' genome. Together, our results show that a mixture of DBD, EC, and Black Queen may operate simultaneously in the human gut microbiome, adding to a growing body of evidence that these eco-evolutionary processes are key drivers of biodiversity and ecosystem function.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}